








Servicios Personalizados
Revista

Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiência e Cultura
versión impresa ISSN 0009-6725versión On-line ISSN 2317-6660
Cienc. Cult. v.62 n.spe1 São Paulo 2010

Biologia da flôr do cafeeiro Coffea arabica L. (*)
ALCIDES CARVALHO e C. A. KRUG
Instituto Agronômico de Campinas. S.P.
Quando se organizou, em 1932, o plano geral de trabalhos com o cafeeiro na Secção de Genética do Instituto Agronômico de Campinas, incluiu-se, como um dos itens básicos, d estudo da biologia da flor da espécie C. arabica, que é a mais cultivada em São Pauto. O conhecimento dêste assunto era de interêsse, não só para se delinear os métodos de polinização artificial, como para verificar se essa espécie era autógama ou, até que ponto, de fecundação cruzada. Da solução dêsse problema também dependia a escolha dos processos mais adequados de melhoramento.
Poucos dados se encontravam na literatura. Os existentes, apresentados por autores holandeses e alemães, se referiam mais à espécie Coffea canephora, reconhecidamente autoestéril e de fecundação cruzada (1, 2, 8,). Taschdjian (7), que por algum tempo trabalhou no Instituto Agronômico de Campinas, fez várias observações sôbre a biologia da flôr da espécie arabica. Usando pela primeira vez, em café, um indicador genético no estudo da polinização estranha (3), isto é, o característico recessivo purpurascens, chegou à conclusão de que cêrca de 50% das mudas obtidas, em viveiro, eram resultantes da polinização cruzada natural. Concluiu que o cafeeiro arabica era planta essencialmente alógama.
No início dos atuais trabalhos sôbre o melhoramento do café, a partir de 1932, nova série de observações sôbre a biologia da flôr do C. arabica foi feita por C. A. Krug e J. E. T, Mendes. Assim é que se determinou o momento da abertura do botão floral, da dehiscência das anteras, a quantidade de pólen produzido, os agentes da polinização, além de novos dados sôbre a porcentagem de fecundação cruzada natural. Usou-se, para êstes estudos, o mesmo indicador genético de Taschdjian, tendo-se chegado a conclusões semelhantes (5, 6).
Não sendo porém, suficientes os dados até então obtidos, resolveu-se, em 1945, esboçar um novo e detalhado plano de estudos sôbre a biologia da flôr, destacando-se, entre outros pontos a serem determinados, o mecanismo da transmissão do pólen nas flôres de C. arabica. Êstes estudos foram realizados na Estação Experimental Central de Campinas e têm por finalidade esclarecer, com a maior aproximação possível, qual a contribuição dos seguintes fatôres na polinização geral nessa espécie de café:
1. Polinização própria — sem o concurso de outros agentes
2. Vento na autopolinização — dentro de uma. flôr e dentro da planta
3. Insetos na autopolinização — dentro de uma flôr e dentro da planta
4. Gravidade na autopolinização — dentro de uma flôr e dentro da planta
5. Vento na polinização estranha
6. Insetos na polinização estranha.
7. Gravidade na polinização estranha.
A determinação do papel dêsses agentes, tanto isolados como em conjunto, é difícil, tendo sido necessário lançar-se mão de artifícios para a obtenção de alguns dos dados em questão. Os resultados obtidos, embora apenas aproximados, contribuem para melhor esclarecer o mecanismo geral da polinização nas flores do café.
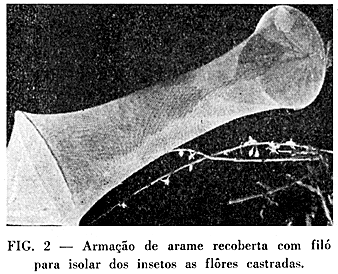
TÉCNICA ADOTADA — Para se determinar o efeito dêstes diferentes fatores, separadamente ou em conjunto, foram usadas técnicas especiais tanto de castração dos botões florais, como do isolamento do cafeeiro e das flôres. Sempre que se precisou isolar a planta do cafezal, usaram-se cafeeiros em barrica, que foram levados ao local mais apropriado. Para se obter a autopolinização própria, isto é, a autopolinização sem o concurso do vento, dos insetos e. da gravidade, foram usados ramos com um ou vários botões por nó, Êstes foram protegidos com uma armação de arame recoberta de uma camada dupla de papel impermeavel. As extremidades dessa armação e a base dos ramos foram firmemente amarradas em estacas de bambú, para impedir a movimentação do ramo no interior. Empregou-se também algodão na extremidade da armação, prêsa à base do ramo, para impedir a entrada de insetos (Fig. 1). Para isolar dos insetos, as flôres castradas ou normais, foram protegidas com armação de arame recoberta de filó (Fig. 2). Ao se estudar o efeito dos insetos na autopolinização ou polinização cruzada, um novo tipo de castração foi adotado, isto é, castração sem eliminar a corola. As flôres assim castradas abrem-se normalmente, porém sem os estames. Para se obter a capacidade total do vento, insetos e gravidade na polinização estranha, usaram-se ramos inferiores de certos cafeeiros no cafezal, cujas flôres foram castradas pelo processo usual (castração com eliminação do tubo da corola e estames (4) (Fig. 3). Empregando-se indicador genético apropriado, pode-se separar as sementes resultantes de polinização estranha daquelas que foram autopolinizadas. Combinações dessas e de outras técnicas foram usadas nas determinações dos efeitos dos agentes isoladamente, combinados dois a dois ou em conjunto.


RESULTADOS OBTIDOS — Os resultados obtidos, num total de 25 itens, repetidos em vários cafeeiros e durante três anos seguidos (1945 a 1947). foram reunidos a fim de indicar a capacidade «máxima» dêsses agentes da polinização e os seus efeitos em flôres normais, isto é, em flôres que não sofreram castração. Como era de se esperar, os dados obtidos nesses dois grupos foram bem diferentes. E' que, na obtenção do efeito «máximo» de cada um dos agentes, empregavam-se vários artifícios, ao passo que, no segundo grupo, usaram-se plantas no cafezal e flôres normais não castradas. Conquanto seja interessante saber qual a contribuição «máxima» do vento, dos insetos e da gravidade na autopolinização e na polinização estranha, compreende-se que se afigura de maior importância prática a determinação do papel que êsses agentes desempenham em flôres normais, nos cafezais. Os resultados obtidos para êsses agentes, em flôres norrnais foram os seguintes:
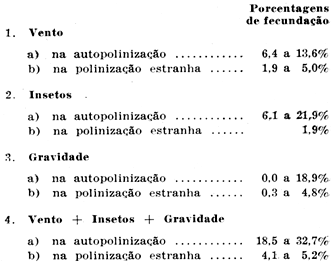
Êsses algarismos referem-se à porcentagem de fecundação, isto é, à porcentagem de sementes formadas em relação ao número inicial de óvulos. Este foi obtido multiplicando por dois o número de flôres utilizadas e adicionando a êsse total o número de frutos com três lojas e a metade do número de sementes concha (sementes que se formam em geral em número de duas por loja). Achou-se melhor utilizar a porcentagem de fecundação em lugar da porcentagem de frutificação, pois esta não levaria em conta os frutos moca que, encerrando uma só semente, iriam exagerar o efeito de alguns agentes.
Vê-se que as porcentagens de fecundação correspondentes à polinização estranha são sempre inferiores às porcentagens na autofecundação. Nota-se ainda que o vento e os insetos têm, aproximadamente o mesmo efeito na autopolinização, sendo menor e mais variável o efeito da gravidade. Quanto à polinização estranha, os dados indicam que o vento parece ser o agente principal. Calculou-se também, para todos os itens, a porcentagem de sementes resultantes da polinização estranha, em flôres normais e no cafezal, em relação ao número de sementes colhidas. Para o efeito conjunto do vento, dos insetos e da gravidade, essa porcentagem variou de 7,3 a 9%. Êsse resultado indica que num cafezal é essa, aproximadamente, a porcentagem de hibridação natural, sendo, portanto, bem mais baixa do que a encontrada por Taschdjian e, anteriormente, por Krug e Mendes, no Instituto Agronômico, À diferença parece relacionar-se com os indicadores genéticos usados em um e outro caso. O primeiro indicador, isto é, o purpurascens, é recessivo em relação ao verde e bronze das folhas novas e suas sementes, em geral, não germinam bem. Além disso, produz menor número de flôres do que as variedades comuns de café as suas pétalas sendo rosadas. O indicador genético cera, usado nos estudos atuais, se assemelha mais à var. typica, dela diferindo apenas na coloração amarelada da semente. Êste característico é também recessivo e devido a um par de fatores principais, Do cruzamento de cera com uma planta normal de sementes verdes, já a própria semente híbrida é verde, mostrando, portanto, o fenômeno de xenia.
Num cafeeiro cera, localizado no cafezal e rodeado de plantas normais, todas as sementes verdes por êle produzidas, representam o resultado da polinização estranha pelos diversos agentes mencionados.
Os dados obtidos nos diferentes cafeeiros utilizados e nos três anos em questão, variaram bastante. Nem podia ser de outro modo, porquanto a ocorrência, por exemplo, de chuvas no dia do florescimento, contribuí para diminuir a ação do vento e dos insetos e os ventos fortes também prejudicam a ação dos insetos, transportando, porém, para mais longe o pólen.
E' de interêsse realçar que foi o conhecimento do mecanismo hereditário do característico cera que permitiu o uso dessa mutação para êstes estudos, que por certo seriam mais difíceis, menos precisos, e mais dispendiosos, se tivessem de ser efetuados com auxílio de outros indicadores genéticos do café.
BIBLIOGRAFIA
1. Cramer, J. S. and P. C. van der Wolk. Em Furwirth, C. Handbuch der landwirtschaftlichen Pflanzenzüchtung. Kaffee. 5:143-161. 1933.
2. Ferwerda, F. P. — A Fazenda. 43:32-35. 1948.
3. Heribert-Nilsson, N. — Zeitschrift für Pflanzenzüchtung. 5: 89-114. 1917.
4. Krug, C. A. — J. of Heredity 26: 325-330, Figs. 14-18. 1935.
5. Krug, C. A. — Revista da Superintendência das Serviços do Café. 20; 863-872. 1945.
6. Krug, C. A. e A. S. Costa — A Fazenda. 42: 35 e 46 a 47. 1947.
7. Taschdjian, E. — Zeitschrift für Züchtung, 17: 341-354. 1932.
8. Zimmermann, A. — Em Kaffee. Auslandbücherei, Berlin. 1928.
(*) Trabalho apresentado à II Semana de Genética, em Piracicaba, em fevereiro de 1949.