









Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO
Compartilhar

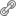 Permalink
PermalinkCiência e Cultura
versão impressa ISSN 0009-6725
Cienc. Cult. vol.71 no.3 São Paulo jul./set. 2019
http://dx.doi.org/10.21800/2317-66602019000300014
ARTIGOS
ENSAIOS
Humanos e formigas utilizam as plantas que proliferaram com a fragmentação da mata atlântica nordestina
Elâine Maria dos Santos RibeiroI; Maria Joana SpechtII; Marcelo TabarelliIII; Bráulio Almeida SantosIV; Pille GerholdV; Rainer WirthVI; Inara Roberta LealVII
IProfessora do curso de licenciatura em ciências biológicas da Universidade de Pernambuco (UPE), campus Petrolina. Email: elaine.ribeiro@upe.br
IIUniversidade Federal de Pernambuco (UFPE).E-mail: joanaspecht@gmail.com
IIIProfessor do Departamento de Botânica da Universidade Federal de Pernambuco (UFPE), credenciado no Programa de Pós-Graduação em biologia vegetal da UFPE. E-mail: mtrelli@ufpe.br
IVProfessor adjunto da Universidade Federal da Paraíba (UFPA), campus João Pessoa, onde coordena o Programa de Pós-Graduação em Ciências Biológicas (Zoologia) e o Laboratório de Ecologia Aplicada e Conservação (LEAC). E-mail: braulio@dse.ufpb.br
VPesquisadora sênior do Departamento de Zoologia, Instituto de Ecologia e Ciências da Terra, da Universidadede Tartu, na Estônia. E-mail: pille.gerhold@ut.ee
VIPesquisador, docente e chefe do Departamento de Ecologia e Sistemática de Plantas da Universidade de Kaiserslautern, na Alemanha. E-mail: with@rhrk.uni-kl.de
VIIProfessora do Departamento de Botânica da Universidade Federal de Pernambuco (UFPE), credenciada nos Programas de Pós-Graduação em Biologia Vegetal e Biologia Animal da instituição. E-mail: irleal@ufpe.br
É sabido que muitas espécies têm suas populações reduzidas e várias se tornam extintas devido à fragmentação das florestas. Como as populações que utilizam as plantas em florestas fragmentadas reagem à modificação na composição de espécies? Neste estudo buscamos entender o porquê de as formigas e os seres humanos compartilharem 50% das espécies de plantas de sua dieta em uma paisagem fragmentada da Mata Atlântica.
Embora muitos dos efeitos da perda e fragmentação de habitats sobre as florestas tropicais já sejam bem conhecidos, ainda existe muito a ser elucidado sobre esse processo. Uma das consequências diretas da perda e fragmentação das florestas é a simplificação dos habitats, através da homogeneização da composição de espécies [1, 2]. Quando estamos tratando das plantas, por exemplo, sabemos que, com a perda e fragmentação das florestas, há predomínio de espécies bem adaptadas às novas condições do habitat (e.g. fragmentos florestais pequenos e isolados, com muita entrada de luz e vento) tais como espécies de crescimento rápido, com muitas sementes pequenas que se auto dispersam ou que dependem de polinizadores e dispersores generalistas. Essas mudanças na flora, base da teia trófica dos ecossistemas terrestres, pode trazer consequências indiretas no padrão de utilização dos recursos vegetais de diversos herbívoros, desde formigas cortadeiras que coletam fragmentos de folhas para cultivar o jardim de fungos do qual se alimentam até humanos que podem utilizar as plantas como fonte de lenha. Mas será que seres tão diferentes como humanos e formigas cortadeiras respondem da mesma forma à fragmentação da floresta com relação à seleção das plantas que utilizam?
A Mata Atlântica é uma dessas florestas tropicais megadiversas que sofreu forte perda e fragmentação, mas que ainda possui uma vasta diversidade de plantas (cerca de 17,7 mil espécies) [2, 3, 4]. Essa diversidade em número de espécies é expressa, por exemplo, pelos tipos de folhas, cujas dimensões podem variar desde o tamanho da cabeça de um alfinete, como os folíolos das folhas partidas do visgueiro, até as folhas da embaúba, cujas dimensões são maiores que um prato. Da mesma forma, essas espécies são diversificadas em relação a outras características, como a composição química, espessura, dureza, presença de pelos e espinhos nas folhas, densidade da madeira e abundância de seus indivíduos na floresta. Assim, cada espécie possui um conjunto de características que influencia desde seu crescimento até o quanto de carbono a espécie vai armazenar em galhos, folhas e troncos. Por exemplo, espécies de folhas grandes e macias têm maior capacidade de estocar carbono atmosférico, o que pode resultar em maiores taxas de crescimento e reprodução da planta. Por outro lado, espécies com folhas grossas e duras podem evitar os danos causados por herbívoros quando comparadas às folhas finas e macias, crescendo mais lentamente e se reproduzindo menos.
Estudos de etnobotânica (ciência que estuda a relação dos seres humanos com as plantas) têm mostrado que populações humanas rurais que fazem o uso direto de plantas selecionam espécies florestais para diversos propósitos, tais como medicinal, construção civil e como fonte de energia (lenha) [3, 5]. Essa seleção das plantas é realizada de acordo com o tipo do uso. Por exemplo, para construção civil e lenha, a escolha é baseada, principalmente, na qualidade da madeira: quanto mais densa (ou dura) a madeira, mais adequada para esse fim [6]. Para uso medicinal, a seleção é feita por meio da análise de propriedades químicas presentes em alguma parte da planta ou do relato de sucesso no uso [7].
Porém, quando se trata de minúsculos insetos sociais, como as formigas cortadeiras do gênero Atta, será que elas também selecionam as plantas que melhor se adequam ao crescimento do fungo que cultivam em suas colônias e do qual se alimentam? Vários estudos enfocaram a seleção de plantas cortadas por formigas cortadeiras e, sim, elas também escolhem não apenas entre espécies diferentes de plantas, mas também entre indivíduos diferentes da mesma espécie e entre partes diferentes da mesma planta [8, 9]. Por exemplo, espécies de plantas pioneiras que crescem rapidamente quando expostas ao Sol são preferidas em relação às plantas que crescem lentamente na sombra, porque as pioneiras têm menos defesas químicas (e.g. terpenos) e físicas (e.g. pelos) contra herbívoros e são mais palatáveis para as formigas [8]. Da mesma forma, folhas murchas são preferidas em relação às não murchas, porque são mais fáceis de cortar e mais palatáveis, bem como folhas jovens são preferidas em relação às velhas porque são mais macias e palatáveis [9].
Na Mata Atlântica nordestina localizada ao norte do rio São Francisco, o setor mais degradado de toda a Mata Atlântica brasileira [1], estudos sobre o uso de plantas como combustível doméstico e de comportamento de forrageamento de formigas cortadeiras foram realizados entre os anos de 2000 e 2013 [3, 8]. Esses estudos foram conduzidos em fragmentos de floresta e seus entornos em uma paisagem rural. Através de entrevistas e caminhadas na floresta com habitantes locais, foi possível identificar as espécies de árvores mais utilizadas para combustível doméstico por pessoas que vivem ao redor dos fragmentos. Para descobrir as espécies utilizadas pelas formigas, foram coletados pedaços de folhas das espécies vegetais presentes nas trilhas das formigas cortadeiras, os quais foram identificados posteriormente no nível de espécie. Embora esses estudos tivessem sido destinados a responder perguntas isoladas para cada grupo biológico (humanos e formigas cortadeiras), os pesquisadores ficaram intrigados com o fato de que 50% das plantas utilizadas como lenha pelas comunidades rurais faziam parte da dieta das formigas cortadeiras (Figura 1). Isso não era esperado porque, de acordo com a teoria existente até então [6, 10, 11], os grupos selecionariam plantas com características opostas: os humanos preferindo plantas de sombra, com crescimento lento, madeira dura, folhas pequenas, espessas, duras e bem defendidas contra herbívoros (plantas características das florestas conservadas), e as formigas cortadeiras preferindo plantas de sol, com crescimento rápido, madeira mole, folhas grandes, finas, macias e pouco defendidas contra herbívoros (plantas pioneiras características das florestas fragmentadas).
Quais seriam os motivos que levaram à seleção em comum de espécies vegetais por humanos e formigas cortadeiras? Na ecologia, a teoria do forrageamento ótimo [12] prediz uma relação de custo/benefício envolvida na escolha de um recurso, ou seja, os organismos selecionam recursos que forneçam mais energia do que a gasta para obtê-los. Com base nessa teoria, não valeria a pena buscar uma madeira mais dura para uso como lenha ou uma planta com folhas mais macias para cultivar fungos se essas são muito raras na floresta. Outra teoria ecológica, a hipótese da aparência [13] prediz que as plantas mais suscetíveis ao ataque de herbívoros são aquelas mais visíveis, ou seja, mais abundantes. A hipótese da aparência tem sido adotada recentemente também em estudos etnobotânicos que defendem que o uso por humanos é afetado pela disponibilidade do recurso [7]. Como espécies de madeira dura estão cada vez mais raras em florestas fragmentadas, é possível que os humanos estejam usando as espécies que são mais abundantes atualmente, ou seja, aquelas plantas pioneiras de crescimento rápido, velhas conhecidas das formigas cortadeiras e que tendem a responder positivamente à fragmentação da floresta. De fato, estudos na Mata Atlântica e na Floresta Amazônica [14, 15] têm demonstrado que a perda de habitats gera a proliferação de espécies que se adaptam às novas condições microclimáticas como, por exemplo, maior incidência luminosa e mais vento.
Para verificar essa hipótese, primeiro foi avaliado se a densidade (dureza) da madeira influenciava a escolha das espécies pelos moradores que vivem próximo às florestas estudadas. Diferentemente do relatado na literatura [6], constatou-se que as espécies mais usadas não são as que possuem maior densidade de madeira, não sendo, portanto, essa característica uma boa descritora do padrão de uso das espécies como combustível. Posteriormente, utilizando a abundância de indivíduos das espécies de plantas que ocorrem nos fragmentos de floresta como um descritor da sua visibilidade, foi avaliado se as espécies utilizadas como lenha eram as mais abundantes, o que foi corroborado, ou seja, os moradores usavam as espécies mais abundantes presentes nos fragmentos de floresta ainda existentes em sua paisagem rural (Figura 1). Apesar dessas espécies não serem as de madeira mais dura, sua densidade ficou em torno de 0.65 g/cm3, valor similar ao de outras espécies de uso comercial, como Eucalyptus, que tem densidade de madeira em torno de 0.61 g/cm3. Por fim, além da abundância, a distância às plantas utilizadas foi um fator importante na decisão dos moradores, sendo as espécies mais próximas às bordas da floresta mais usadas, devido à dificuldade de carregar o peso da madeira até os domicílios.
Quanto às formigas cortadeiras, como previsto e relatado na literatura, foram selecionadas espécies de plantas com folhas mais macias e, também, mais abundantes, uma vez que as plantas pioneiras possuem folhas mais macias e são mais abundantes do que espécies características da floresta madura, especialmente nas bordas (Figura 1). Esses resultados nos indicam que tanto humanos quanto formigas utilizam as espécies mais abundantes nos fragmentos de floresta.
Além de identificar as plantas selecionadas por humanos e formigas cortadeiras, verificamos que as espécies utilizadas eram mais próximas em termos de relação de parentesco, como primos de uma mesma família (Figura 2). As formigas cortadeiras, por exemplo, concentraram seu forrageamento nas plantas com folhas mais macias, característica que ocorre frequentemente em alguns grupos de plantas específicos, como as famílias Euphorbiaceae e Melastomataceae. As espécies com maiores valores de densidade da madeira também formaram grupos filogenéticos específicos (e.g. Sapotaceae e Primulaceae), apesar de tal característica não ter sido levada em conta na seleção pelos moradores dos entornos dos fragmentos da floresta.
O desmatamento e a fragmentação da floresta acabam direcionando o uso das espécies tanto pelas formigas como pelos humanos, forçando seres completamente diferentes a utilizarem as espécies que conseguem proliferar mesmo depois de várias décadas de exploração desordenada. É sabido que muitas espécies têm suas populações reduzidas e várias se tornam extintas por conta da perda e fragmentação de habitats. Todavia, nossos achados reforçam a ideia de que algumas espécies se beneficiam ou apenas se adaptam às novas condições. No caso das populações humanas, a fragmentação das florestas leva a uma migração no padrão de exploração baseada em recursos mais nobres para aqueles recursos que sobram. Esse é o mesmo padrão encontrado em outras formas de exploração de recursos naturais como a pesca e a extração comercial de madeira. Além disso, como observado aqui, é provável que não apenas espécies, mas grupos filogenéticos inteiros sejam perdidos com a fragmentação das florestas. Ainda não é possível estimar os impactos dessa convergência no uso das espécies vegetais, mas alguns cenários são possíveis como a persistência apenas de espécies pioneiras com maior grau de adaptação às áreas fragmentadas, cujas possíveis consequências precisam ser melhor avaliadas.
AGRADECIMENTOS
Os autores agradecem aos revisores anônimos pelas contribuições que possibilitaram a melhoria do trabalho e às instituições que o financiaram (CNPq 540322/01-6, 243000/02, 165867/2015-9, Capes 007/01, DFG WI 1959/1-1, Estonian Research Council - grant PUT1006).
REFERÊNCIAS
1. Lôbo, D.; Leão, T.; Melo, F. P. L.; Santos, A. M. M.; Tabarelli, M. "Forest fragmentation drives Atlantic forest of northeastern Brazil to biotic homogenization", Diversity and Distributions, 17, 287-296. 2011.
2. Tabarelli, M; Aguiar, A. V.; Leal, I. R.; Lopes, A. V. Serra Grande: uma floresta de ideias. Recife: Editora Universitária da UFPE, 2013.
3. Specht, M. J.; Pinto, R. S. S.; Tabarelli, M.; Melo, F. P. L. "Biodiversidade queimada: Uso de lenha para cozinhar ameaça Mata Atlântica do Nordeste", Ciência Hoje, 308, 28-31. 2013.
4. Flora do Brasil 2020 em construção. Jardim Botânico do Rio de Janeiro. Disponível em: <http://floradobrasil.jbrj.gov.br/ Acesso em: 17 Jun. 2018.
5. Anderson, E. N.; Pearsall, D.; Hunn, E.; Tuner, N. Ethnobiology. New Jersey: John Wiley & Sons, 2011.
6. Ramos, M. A.; Medeiros, P. M.; Almeida, A. L. S.; Feliciano, A. L. P.; Albuquerque, U. P. "Can wood quality justify local preferences for firewood in an area of caatinga (dryland) vegetation?", Biomass and Bioenergy, 32, 503-509. 2008.
7. Santos, C. S.; Barros, F. N.; Paula, M.; Rando, J.; Nascimento, V. T.; Medeiros, P. M. M. "What matters when prioritizing a medicinal plant? A study of local criteria for their differential use", Acta Botânica Brasilica, 32, 297-302. 2018.
8. Falcão, P. F.; Pinto, S. R. R.; Wirth, R.; Leal, I. R. "Edge-induced narrowing of dietary diversity in leaf-cutting ants", Bulletin of Entomological Research, 101, 305-311. 2011.
9. Ribeiro-Neto, J. D.; Pinho, B. X.; Meyer, S. T.; Wirth, R.; Leal, I. R. "Drought stress drives intraspecific choice of food plants by Atta leaf-cutting ants", Entomologia Experimentalis et Applicata. 144, 209-215. 2012.
10. Howard, J.J. "Index to Ecology, volume 69, 1988", Ecology, 69, 250-260.1988.
11. Mundim, F. M.; Costa, A. N.; Vasconcelos, H. L. "Leaf nutrient content and host plant selection by leaf-cutter ants, Atta laevigata, in a Neotropical savanna", Entomologia Experimentalis et Applicata, 130, 47-54. 2008.
12. Pyke, G. H.; Pulliam, H. R.; Charnov, E. L. "Optimal foraging: a selective review of theory and tests", Quarterly Review of Biology, 52, 137-154. 1977.
13. Feeny, P. "Plant appearance and chemical defense". In: Biochemical interactions between plants and insects. New York: Plenum. 1976.
14. Oliveira, M. A.; Santos, A. M. M.; Tabarelli, M. "Profound impoverishment of the large-tree stand in a hyper-fragmented landscape of the Atlantic Forest", Forest Ecology and Management, 256, 1910–1917. 2008.
15. Laurance, W. F.; Nascimento, H. E. M.; Laurance, S. G.; Andrade, A. C.; Fearnside, F. M.; Ribeiro, J. E. L.; Capretzr, R. L. "Rain forest fragmentation and the proliferation of successional trees", Ecology, 87, 469–482. 2006.
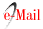
{kind=link}