








Services on Demand
Journal

Article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCiência e Cultura
Print version ISSN 0009-6725On-line version ISSN 2317-6660
Cienc. Cult. vol.62 no.spe1 São Paulo 2010
Enxertias entre plantas de diferentes famílias
K. SILBERSCHMIDT
Instituto Biológico de São Paulo
Virgílio dá, na sua «Georgica», uma florida descrição de macieiras enxertadas sôbre plátanos e de mirtas que continuam vegetando viçosamente sôbre porta-enxertos de carvalhos. Na literatura da antiguidade e da idade média, encontramos exemplos semelhantes, com grande frequência. Assim, o escritor árabe Ibn Wahhschija, citado por E. Bergdolt (1) dá instruções muito detalhadas sôbre a maneira de se enxertar um limoeiro sôbre uma oliveira. Afirma ainda o autor árabe, que o limoeiro assim enxertado produz lindos limões, que, em tamanho, forma, côr e no seu teôr em óleo, lembram azeitonas.
Surge, então, agora, a pergunta, se tais enxertias entre plantas de diversas famílias pertencem apenas ao reinado da lenda e do mito ou se verdadeiramente essas enxertias são viáveis, segundo os modernos conceitos científicos.
Deve-se explicar, desde logo, que, do ponto de vista científico, há grande interêsse na obtenção de enxertias interfamiliares.
Já na sua aplicação a plantas de espécies afins (não interfamiliares) o método de enxertia tem servido para resolver muitas questões da anatomia, bioquímica e até de fitopatologia e é evidente que ainda outras questões interessantes poderiam ser estudadas em enxertias interfamiliares. Para dar uma idéia dos problemas fundamentais que podem ser atacados pelo método da enxertia, vamos mencionar, pelo menos, um exemplo, que se refere à localização, na planta do fumo, do sítio em que se forma o alcaloide nicotina. Durante mais de um século admitiu-se (9) que a fôlha seria o centro principal de formação da nicotina, a qual ali permaneceria sem ser transportada para outros órgãos. Em trabalhos recentes, porém, Dawson (3, 4), estudando a distribuição de nicotina em enxertias de fumo (Nicotiana tabacum L.) sôbre tomate (Sol. lycopersicum L.) e de tomate sôbre fumo, verificou que em condições normais a nicotina é formada nas raizes e de lá transferida para as fôlhas. As plantas de fumo, enxertadas sôbre cavalos de tomate, perderam, nas experiências de Dawson, pouco a pouco o seu teor inicial em nicotina, ao passo que os tomateiros enxertados sôbre cavalos de fumo acumularam nicotina nas suas fôlhas. Dawson tira, destas experiências, a conclusão de que a nicotina é formada apenas nas raizes de fumo e que de lá pode ser transferida até para folhas de uma outra espécie, o tomateiro. Êste trabalho é um ótimo exemplo para mostrar como o método de enxertia pode contribuir para resolver problemas de bioquímica. E' facil imaginar a que resultados interessantes poder-se-ia chegar, se, de fato, dispuzermos de um método, para combinar, numa só enxertia, plantas de qualquer família. Lembro a possibilidade de enxertar Digitalis sôbre fumo, ou Atropa belladona sôbre Cinchona ledgeriana. Mas será que esta possibilidade existe realmente?
Até há muito pouco tempo, os autores que mais se ocuparam dêste problema, não acreditavam que plantas pertencentes a famílias diferentes pudessem formar enxertia, de modo duradouro. Assim escreve Winkler (15, pág. 772) que não se conseguiu, por enquanto, um pegamento duradouro entre representantes de diferentes famílias. Molisch (8, pág. 269) acha que as indicações de autores anteriores que se referem ao sucesso em enxertias interfamiliares, carecem de fundamento. E ainda, em 1948, Nickell (10) constata que até hoje, em geral, se admite que enxertias entre plantas não afins são impossíveis.
Não me parece, porém, que tais opiniões estejam de acôrdo com o estado atual dos nossos conhecimentos e até duvido que elas pudessem ser seriamente defendidas no tempo em que foram pronunciadas. Antes de mencionar argumentos em contrário, devemos, pelo menos em poucas palavras, procurar explicar como pode haver divergência de opiniões entre diversos autores sôbre o sucesso de um mesmo caso de enxertia interfamiliar. Todos os autores, naturalmente, admitem que não faltam, na literatura, referências a tentativas para a obtenção de enxertias entre plantas de diversas famílias e a discussão gira apenas em tôrno do problema, de se saber se o contato estabelecido entre os componentes da enxertia era ou não «duradouro». Para o leigo, a decisão desta questão parece não oferecer qualquer dificuldade. Na realidade, porém, é possível manter enxertias vivas, durante semanas, mesmo sem sinais anatômicos de pegamento, desde que, sob cuidados especiais, sejam guardadas em câmara úmida. Se uma parte dos autores já está inclinada a reclamar sucesso para enxertias dêsse tipo, apenas porque o enxerto, sob os cuidados do jardineiro, se manteve verde durante semanas, os pesquisadores mais severos consideram bem sucedida uma enxertia apenas quando se prova o estabelecimento, na zona de enxertia, de «janelas de contacto» (isto é, áreas em que as células dos dois componentes estão em contacto direto) ou até de sistema condutor completo (xilema e floema). Só então, segundo êsses autores, os dois componentes constituem, de fato, uma nova unidade em estrutura e metabolismo. Por isso, êsses autores exigem que para avaliar o sucesso de uma enxertia, se proceda a um exame anatômico da zona de enxertia. Vejamos agora, quais as indicações da literatura, relativas ao sucesso em enxertias interfamiliares, que satisfazem a êste último critério.
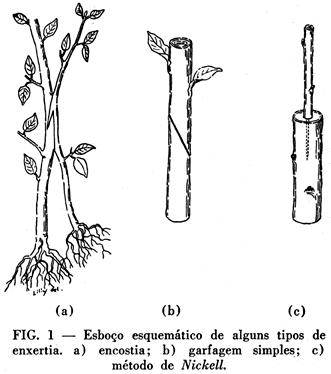
A partir do fim do século passado, o botânico francês Daniel publicou uma série de pequenas comunicações em que, frequentemente, ' faz menção do sucesso obtido em enxertias interfamiliares. Já que as descrições dele são pouco explícitas e não incluem detalhes anatômicos, os casos por êle indicados, de enxertias interfamiliares, não são, geralmente considerados como provados. Achamos, porém, possível — levando em conta os resultados obtidos por outros autores — que êle, pelo menos em alguns casos, de fato, conseguira pegamento entre componentes de enxertia, pertencentes a famílias distintas. Queremos destacar aqui, por exemplo, o caso da enxertia de couve-rábano (Cruciferae) sôbre tomateiro (Solanáceae), cuja descrição (2) nos parece merecer fé. Daniel recomendou, com razão, para a obtenção de enxertias tão delicadas o emprêgo do método de encostia (Fig. 1, a) — no qual, inicialmente, os dois componentes ficam com o seu próprio sistema radicular — e insistiu na necessidade de escolher, para tais enxertias, plantas que se acham na mesma fase de vegetação. Mesmo assim, pode-se compreender, que muitas das afirmações de Daniel, fossem aceitas apenas com uma certa reserva. Já não se justificaria, porém, uma tal atitude com referência aos resultados de Simon (13). Êste autor, enxertando garfos de Solanum melongena L. (Solanaceae) em 24 cavalos de Iresine lindenii (Amaranthaceae), conseguiu, em 6 casos (25%), um verdadeiro pegamento entre os componentes. O exame anatômico, procedido numa enxertia que se manteve viva durante 2 meses e meio, revelou a existência, na zona de enxertia, de verdadeiras «janelas de contacto» e a presença de feixes condutores. Constitue, assim, êste caso de enxertias entre Solanaceae e Amaranthaceae o primeiro exemplo de enxertias interfamiliares que desafiam mesmo uma crítica bastante severa.
A partir daquele ano (1930), avolumaram-se as referências da literatura a casos perfeitamente documentados de enxertias interfamiliares bem sucedidas. A monografia sôbre enxertias do pesquisador russo Krenke (7) traz uma lista de 15 tipos de enxertias entre representantes de diversas famílias. Uma destas enxertias, cuja fotografia se acha no citado livro, e que é representada aqui na nossa figura 2, mostra o tomateiro (Solanaceae) como porta-enxerto e o crisântemo (Compositae) como garfo. Esta enxertia que, originariamente, foi obtida por Gladkov (6), ficou viva durante 5 mêses e o enxerto chegou a produzir numerosas sementes. Numa outra dessas enxertias heteroplásticas obtidas por Gladkov, em que o hiperbionte (= enxêrto) é representado por Artemisia absinthium L. (Compositae) e o hipobionte (= cavalo) pelo tomateiro (Solanaceae) verificou-se a formação de novos feixes condutores na zona de enxertia. Um número relativamente grande de enxertias interfamiliares foi obtido, pouco tempo mais tarde por Silberschmidt (11, 12). Se lançarmos uma vista d'olhos sôbre as fotografias, que acompanham êsses trabalhos, veremos, por exemplo, uma muda de couve (Cruciferae), como hiperbionte, sôbre um porta-enxêrto de fumo (Solanaceae). Esta enxertia, que se manteve viva durante mais de 6 semanas e que foi apenas sacrificada para possibilitar o estudo anatômico da zona de enxertia, apresentou, na medula, algumas «janelas de contacto» bem acentuadas sem que nelas já se pudesse notar a diferenciação de traqueides. Nas «janelas de contato», as células de couve confinavam diretamente com as de fumo, interrompendo, nestes lugares «a camada isoladora», formada por restos de células lesadas pelo canivete do enxertador, camada esta que, em enxertias não pegadas, separa os dois componentes em tôda a extensão do corte. Outras enxertias interfamiliares, cujas ilustrações se acham nos trabalhos de Silberschmidt, apresentam a figueira do inferno (Datura stramonium L., Solanaceae) como cavalo e, como enxêrto, ora o crisântemo e Zinnia (Compositae), ora Impatiens sultani (Balsaminaceae), ou ainda Digitalis, purpurea L. O estado de pegamento de tôdas essas enxertias foi geralmente o mesmo da enxertia já descrita entre couve e fumo. Do ponto de vista da constituição de nova unidade metabólica, a enxertia entre Digitalis e Datura merece certamente ser estudada de novo.

Antes de completar esta pequena lista de referências da literatura, relativa a casos de enxertias bem sucedidas entre representantes de diversas famílias de plantas, queremos explicar quais são os caracteres comuns atribuídos às enxertias interfamiliares pelos autores até agora mencionados. Concordam os autores, principalmente, em admitir que as diferenças entre tais enxertias e aquelas entre plantas mais afins, são graduais e não absolutas. Não duvidam Simon, Krenk e Silberschmidt que, geralmente, o pegamento entre plantas afins seja mais facil e mais rápido. Afirmam porém, que, em certos casos, a falta de afinidade sistemática entre duas plantas não exclue a possibilidade de uni-las, com sucesso, numa enxertia. Concordam também os mencionados autores em seguir, para a obtenção das enxertias interfamiliares, os mesmos métodos que foram desenvolvidos, no curso dos séculos, para conseguir enxertias de plantas afins. Assim, seguindo êste método parece indicado que o câmbio do hiperbionte seja diretamente ajustado com o do hipobionte (Winkler 15, pg. 776). Geralmente, o enxertador também trata de adaptar, exatamente, a casca dos dois componentes de enxertia. Justamente por causa da conveniência de sobrepor tecidos homólogos do enxêrto e cavalo, escolhiam-se, de preferência, sobretudo em se tratando de plantas herbáceas, caules de espessura igual. (Veja o método de enxertia, chamado garfagem simples, representado em fig. 1-b).
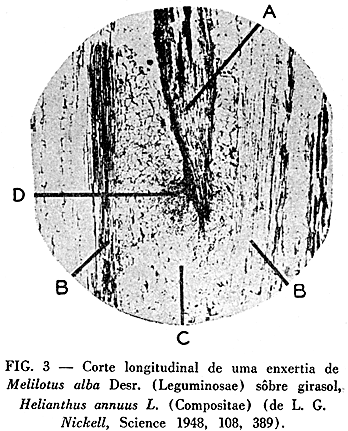
Em muitos dêstes pontos, a recente comunicação de Nickell (10), que também trata da execução de enxertias interfamiliares, representa uma inovação quase revolucionária. O trabalho dá a idéia de que tais enxertias, entre plantas sistematicamente não relacionadas, podem ser conseguidas com a maior facilidade. Segundo êste novo processo o cavaleiro desprovido de fôlhas e a cuja base se dá a forma de bisel, é inserido na medula do porta-enxêrto (Fig. 1-c). Evita-se, expressamente, a justaposição dos câmbios ou dos elementos vasculares dos dois componentes e escolhe-se, geralmente, um cavalo, cuja espessura supera a do enxerto, consideravelmente. Até agora, Nickell conseguiu obter, segundo êste método, enxertias bem pegadas entre Leguminosae como enxêrtos (Melilotus alba e Vigna sinensis Endl.) e Compositae (Helianthus annuus L.) ou Solanaceae (Nicotiana tabacum L.) como cavalos. Em um dos casos, observou-se, na medula do porta-enxêrto, a formação de feixes lenhosos (Fig. 3). Com o aparecimento do trabalho de Nickel, o interêsse dos pesquisadores em enxertias interfamiliares recebeu forte estímulo, pelo menos nos Estados Unidos.
O diretor do Jardim Botânico de Brooklyn (N. Y.), prof. Dr. G. S. Avery convocou, inspirado por êste interêsse, em fins do ano passado um simpósio sôbre enxertia, em que tomaram parte, além do prof. Avery, o prof. R. N. Dawson (Columbia Univ.), o Dr. L. M. Black (Brooklyn Botanical Garden), o Sr. L. G. Nickell (Yale Univ.) e o autor destas linhas, que se achava, então, em viagem de estudos aos Estados Unidos. Discutiram-se, nêsse simpósio, as limitações e as possibilidades futuras para enxertias interfamiliares e principalmente para o processo de Nickell. Frizou-se, que até agora, mesmo com o novo método, poucos casos de pegamento perfeito entre plantas de diversas famílias foram obtidos. Ficou também claro que outros autores além dos mencionados por Nickell, se tinham ocupado com a tentativa de obter enxertias interfamiliares.
Poder-se-ia ainda acrescentar, que também outros autores, como por exemplo, Funck (5), já anteriormente tinham verificado que as alterações anatômicas, induzidas numa enxertia, podem ser estudadas na medula, com a maior facilidade.
Por outro lado, todos os participantes do simpósio reconheceram a grande importância que o método de Nickell poderá adquirir para a obtenção de um número mais vasto de combinações entre representantes de diversas famílias e destacaram principalmente dois terrenos científicos que se beneficiariam consideravelmente com o estudo mais aprofundado, de enxertias interfamiliares.
Um dêstes terrenos seria, sem dúvida, a Genética e o outro a Fitopatologia.
O interesse da genética, nesta questão, é de grande atualidade. Em desacôrdo com opiniões de Daniel que se baseara mais em teoria do que em amplos fundamentos experimentais, já há 30 anos, o botânico Winkler (14), numa monografia sôbre o problema da enxertia, tinha estabelecido, que as influências mútuas entre enxêrto e cavalo possuem o carater de modificações e não alteram as qualidades «específicas» dos componentes. Desde então, esta opinião de que o enxêrto e cavalo conservam, na enxertia, inteiramente os respectivos caracteres genotípicos, foi confirmada pelo exame da descendência de milhares de enxertias. Recentemente, porém, a escola de Lysenko, na Rússia soviética, afirma ser o processo de enxertia um dos fatores mais eficientes para induzir experimentalmente alterações das qualidades genéticas. O material experimental apresentado até hoje, em favor desta opinião (que aliás aos conhecedores da literatura e especialmente dos trabalhos de Daniel não parece tão «nova») é tão pouco convincente que não encontrou nenhum apôio da parte dos geneticistas mais abalizados. Até agora, ambos os lados, nesta disputa, basearam-se largamente nos resultados obtidos com enxertias entre plantas relativamente afins. Talvez, a obtenção no futuro de um número vasto de enxertias interfamiliares forneça um material bastante interessante para estudar novas facetas dêste problema.
Finalmente foi, no referido simpósio, também lembrada a grande vantagem que o fitopatologista, principalmente o virologista, poderá tirar do novo processo. Há, entre as doenças de virus vegetais, muitas, que apenas por enxertia podem ser transmitidas a outras plantas. Para conhecer as espécies hospedeiras de um virus, é necessário incluir na experimentação grande número de «eventuais» espécies susceptíveis. Por enquanto, êstes ensaios de transmissão, com tipos de virus apenas transmissíveis por enxertia, tinham que se limitar largamente a espécies dentro de uma só família de plantas. Foi então, durante o simpósio, manifestada a esperança de que futuramente será mais facil transmitir doenças de virus, experimentalmente, por enxertias, a espécies taxonomicamente bem, distantes daquela em que essas doenças ocorrem espontaneamente.
Mas até chegarmos a êste ponto, muito trabalho resta a fazer e principalmente deve ser estudada a questão, se na realidade, o novo método de enxertia oferece vantagens para a obtenção de enxertias interfamiliars. Alguns ensaios comparativos sôbre esta questão, já estão em andamento, na Secção de Fisiologia Vegetal do Instituto Biológico, em São Paulo.
Mas em todo caso já se pode afirmar hoje, com segurança, que enxertias bem sucedidas entre plantas pertencentes a diversas famílias não existem apenas no reinado da fantasia, mas podem pelo menos em alguns casos, ser obtidos, embora com certas dificuldades técnicas, com os métodos desenvolvidos na experimentação científica.
ABSTRACT
Reference is made to the papers of various authors (Daniel L., Simon S., Krenk N. P., Silberschmidt K.), who studied the behavior of graftings between plants of different families. Recently in the United States a new procedure for obtaining such graftings has been described by L. G. Nickell. The author, during a recent trip to the U. S., took part in a «symposium», held in the Brooklyn Botanical Garden, with the purpose to discuss the possibility of making use, in a near future, of interfamily graftings for the solution of some problems of biochemistry, genetics and virus research.
BIBLIOGRAFIA
1. BERGDOLT, E. — 1934. Zur Geschichte der Botanik im Orient II. Über einige Pfropfungen. Ber. d. deutsch. Bot. Ges. 52:87-94.
2. DANIEL, L. — 1910. Sur les variations spécifiques du chimisme et de la structure provoquées par le greffage de la Tomate et du Chou cabus. C. r. Acad. Sci. Paris 162, 1910.
3. DAWSON, R. F. — 1941. The localization of the nicotine synthetic mechanism in the tobacco plant. Science 94, 396-397.
4. DAWSON, R. F. — 1942. Accumulation of nicotine in reciprocai grafts of tomato and tobacco. Ann. Journ. of Bot, 29, 66-71.
5. FUNCK, R. – 1929. Untersuchungen über heteroplastische Transplantationen bei Sonaceen und Cactaceen. Beitr. Biol. Pflanzen 17:404-468.
6. GLADKOV, W. S. — 1933. Beiträge zur Frage der Pfropfungen zwischen Pflanzen verschiedener Familien. Phaenogenetische Variabilitaet, Bd. II Timiriaseff - Institut f. Biologie, Moskau, 289.
7. KRENKE, N. P. — 1933. Wundkompensation, Transplantation und Chimären bei Pflanzen. Berlin, Julius Springer 1933, 934 pg.
8. MOLISCH, H. — 1930. Pflanzenphysiologie als Theorie der Gärtnerei. Jena, Gustav Fischer, 368 pg.
9. MOTHES, K. — 1930. Das Nikotin im Stoff wechsel der Tabakpflanze. Apothekerzeitung Nº 13, 1-6.
10. NICKELL, L. G. — 1948. Heteroplastic grafts. Science 108: 389-390.
11. SILBERSCHMIDT, K. — 1935. Die Abhängigkeit des Pfropferfolges von der systematischen Verwandtschaft Partner. 7.sch. f. Bot. 29: 65-137.
12. SILBERSCHMIDT, K. — 1936. A importância da «especie» no systema das plantas e a tentativa da sua delimitação por novos methodos. Archivos do Instituto Biológico 7: 33-50.
13. SIMON, S. V. — 1930. Transplantations-versuche zwischen Solanum melongena und Iresine Lindeni. Jb. wiss. Bot. 72: 137-160.
14. WINKLER, H. — 1912. Untersuchungen über Pfropfbastarde. Teil 1, Jena 1912.
15. WINKLER, H. — 1924. Die Methoden der Pfropfung bei Pflanzen. Abderhalden's Hb. d. biol. Arbeitsmethoden. Abt. XI, 2, 766-798.